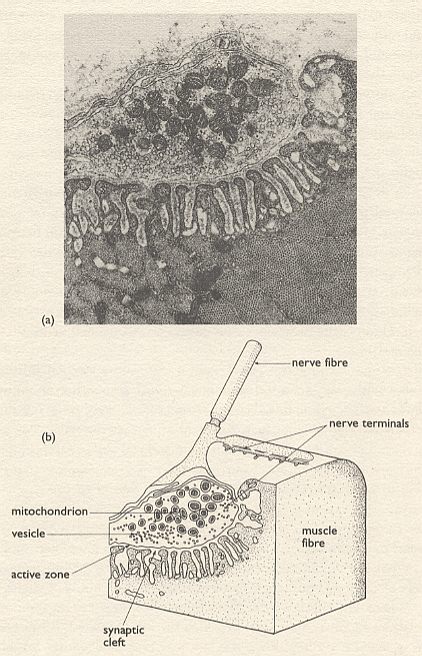
In this and the following note much is about the nature of the junction (synapse) between nerve fiber and muscle cell. Before starting these notes, we here insert an electron micrograph and a diagram of such a junction and some information. The discussion in the notes is about whether such a synapse is chemically continuous or not.
In addition to physiologists, biochemists can (in their way) describe in exquisite detail the processes that go on when a signal (an action potential) passes down a nerve fibre. The potential depends on the lipid and protein structure of the nerve cell membrane, on the differential distribution across the membrane of the ions of potassium (K) and sodium (Na), on the ubiquitous ATP, and so on. At the junction between nerve and muscle (the synapse), defined in considerable anatomical detail, there are tiny membrane-bound packages (vesicles) containing transmitter molecules which can be released when the action potential arrives at the synapse, [these transmitter molecules] diffuse across to the muscle, and there begin the biochemical cascade that ultimately results in the actin and myosin filaments sliding past one another (= muscle twitch). The process is summed up in the above Figure.
In the appearance of cholinesterase [an enzyme which splits, and hence inactivates, acetylcholine (itself being a neurotransmitter, chemically passing on the impulse of a nerve fiber)] at the "transition from nerve impulses to muscles" we do not see a process consisting of replacement of, or speaking against, the bonding relation, but a supporting functional mechanism, a chemism working for the inner environment, a chemism put into service or being exploited, because the nervous impulse (close to being a microphysical phenomenon) all by itself could not result in a gross-macrophysical muscle contraction anyhow.
During sensory experience (having some sensible impression from the outside world), lasting a certain amount of time, one can demonstrate, in the relevant conduction nerves, a series of electrical pulses with constant potential. It is the frequency that fluctuates, whose increase is expressed by the intensification of the experience. This constancy of electrical potential during sensory stimulus conduction, and intensification taking place merely as a result of frequency increase, points to the existence of such firm relationships that can only be accomplished by a fixed chemical constitution. Systems [of molecules] may also be capable of an increase, but as a result of entirely different mechanisms which might be summarized, for example, as "increase of production" [more elements, each one not strengthened, being involved.].
In a nerve bundle there often are nerve fibers having totally different conduction velocities. For this one may, without too much trouble, make up complex systems. One may, however, spare oneself this "making up" and simply operate with the datum, evident in the whole remaining body, namely the continuous chemically bonding structure, which may vary, as expressed in the, for example, different stimulus conduction velocities. Better, of course, is the reverse procedure of thinking : deriving from the phenomenon [of different velocities] the simplest and most certain causation. And there one encounters the differences of bonding [causing the differences of velocities], the continuity of which [bonding] gives the Unimol picture. [So difference in chemical bonding in different nerve fibres cause difference of stimulus conduction velocity.]
Also the presence of a stimulus threshold -- (physiologically it is in all cases favorable to exclude uncertainties [for the organism] and not rendering the organism the plaything of coincidences. But it also must have a constitutive reason) -- much better fits the unimolecular bonding than fitting the electrolyte system, which is "good" for all values, also the smallest.